| Brain Research Group >> Research >> Change-point analysis ... | << previous next >> |
Change-points themselves say little about brain functioning; further data processing is necessary for the extraction of useful information in the form of various quantitative indices. The most simple approach is comparing the number of change-points per time unit in EEG obtained from different subjects, or from the same subject in different states, or from the same subject's different brain sites (in different electrode locations). It seems to be natural to suggest that the higher is the number of change-points, the more complex is the EEG structure.
However, the number of change-points is not a robust index; it may be sensitive to various factors, even not related to the brain activity. The probability of accepting a statistically justified decision about the presence of a change-point in an interval of a given duration, for instance, depends on the amount of available information, which, in its turn, depends on sampling (digitizing) rate and on the specific features of the process under study (the higher is the frequency of a periodic process, the more information it can carry). Moreover, it must be taken into account that the dynamics of the electrical potential on an EEG electrode can result from superposition of the activity of different systems, or, more precisely, of different "generators", the neuronal networks generating the electrical potential, each network with its own dynamics of the potential. The functional interpretation of a change-point in a signal produced by a single "generator" can be rather clear, but if there is a superposition of potential dynamics from different signal generators, then the interpretation becomes much more complicated. The number of change-points in this case may vary with changes in the ratio of the overall power of the different generators, if they produce signals with different complexity of the segmental structure. For example, an increase of the relative contribution from a generator producing a low structured signal will cause "blurring" the change-points contributed from a generator with highly structured signal, decreasing the total number of statistically detectable change-points; and vice versa. A relatively low number of change-points should be expected also in a signal resulting from superposition of a number of signals with high number of change-points and roughly the same overall power. Nevertheless, we believe that, with caution and with taking into account other indices, the index of the number of change-points can be used in the analysis of the EEG.
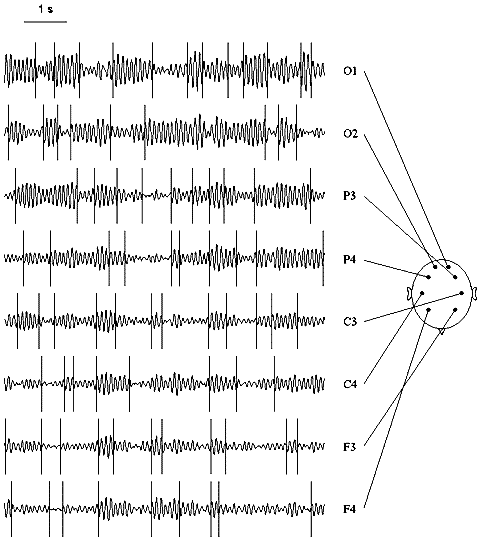
Fig. 7.7. Multi-channel EEG: change-points in alpha activity
The EEG was filtered with bandpass 7.5--12.5 Hz. Change-points (vertical lines) were detected in the basic diagnostic sequence. Subject tw03, eyes closed. Horizontal scale: 1 s.
Variations of the electrical activity across cortical sites can be seen from Fig. 7.7. The overall alpha band power increase from frontal to occipital sites, a pattern usually found in healthy persons. As for change-points, no systematic variations in their number seem to be detectable by visual inspection. Quantitative analysis of this index, however, reveal some regularities. In Fig. 7.8, variations of the number of change-points across subjects are evident. For instance, the number of change-points was especially high for subject tw09 and low in subjects tw07 and tw08 (in the latter case, the alpha rhythm was high but poorly modulated). A certain similarity was found in identical twin pairs (tw07 and tw08 was such a pair), which is in agreement with the other authors' data about the genetic determination of at least some of the EEG characteristics.
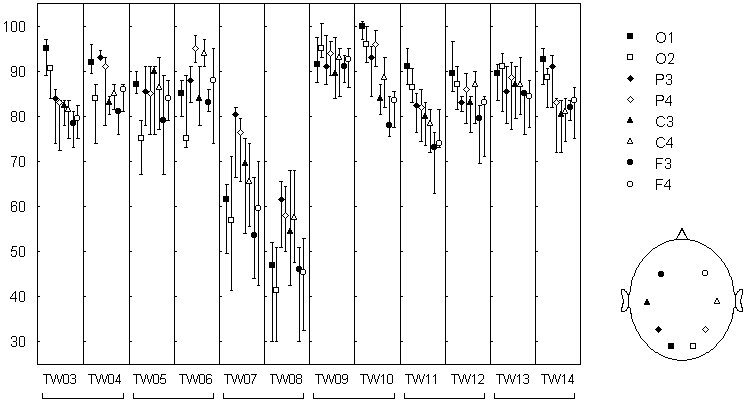
Fig. 7.8. Rate of alpha power change-point occurence in different EEG channels
Genetically identical twins (twin pairs: tw03--tw04, tw05--tw06, etc.). The EEG was recorded in eyes closed condition with 8 electrodes at standard positions (O1, O2, P3, P4, C3, C4, F3, F4). Each EEG channel was filtered with bandpass 7.5--12.5 Hz (alpha). The change-points were detected in the basic diagnostic sequence. Number of change-points in 1 minute EEG (median +-25%; n=3...8 for tw03-tw07; n=10 for tw08-tw14).
The most important finding was the dependence of the number of change-points on the electrode location. In most of the subjects a frontal-occipital gradient was found: the number of change-points was highest in occipital areas (O1 and O2) and lowest in frontal areas (F3 and F4). This dependence did not coincided with the dependence on the electrode location for the EEG pattern and, in particular, for the alpha band power: for example, in tw09 the gradient was almost absent for the change-point number while was well-defined for the alpha band power; his twin brother tw10 had the same gradient for the power and a very clear gradient for the change-point number. Consequently, the gradient of the change-point number hardly was just a reflection of the power gradient, but could be determined by some "structural" features of the alpha activity dynamics, which vary across cortical areas and across subjects.
This view is consistent with a large body of data, obtained with a broad variety of methodical approaches and suggesting the existence of a number of "generators" of alpha activity, which occupy different cortical areas and produce alpha activity with different dynamical characteristics (e.g., Lehmann 1971; Thatcher et al. 1986; Ozaki & Suzuki 1987; Lutzenberger 1997; Florian et al. 1998). Though the structural characteristics of the EEG are still poorly understood, there are strong grounds to believe that they differ for the different "alpha generators" (Basar & Schurmann 1997; Basar et al. 1997; Lutzenberger 1997).
The data presented in Fig. 7.8 were obtained in rest condition with eyes closed. In the rest condition but with eyes open, the alpha band power decreased and the pattern of alpha activity substantially altered, but the number of change-points did not differ significantly for any electrode location (significance level p>0.05, Wilcoxon matched pairs test). Our data still not allow to decide whether the lack of significant difference came from the stability of individual structural characteristics of EEG alpha activity, or the method was simply not quite sensitive to such characteristics. A situation may be rather complex: for example, in eyes open condition the contribution of the high alpha segments to the EEG may be reduced, then the signal to noise ratio will grow for "finer" change-points, resulting in the absence of detectable difference in the total number of change-points. In any case, the data shown in Fig. 7.8 demonstrate that the index of the number of change-points possesses sensitivity to some EEG features, since it varies across subjects and brain site.