| Brain Research Group >> Research >> Change-point analysis ... | << previous next >> |
The change-points, as it can be seen in Fig. 7.9, often appear near-simultaneously in more than one channel. It is naturally to treat the change-points appearing close in time in a number of EEG channels as a single complex. We will call it a change-point synchrocomplex, or simply synchrocomplex. Since the different EEG channels represent the activity from different cortical areas, which lies under the corresponding electrodes, the synchrocomplex represents the topography of sharp transformations of electric activity, which occur in these areas in synchrony or almost in synchrony and probably reflect the functional connectiveness of these areas.
The spatial configuration of the functionally connected areas may vary dramatically within even short time intervals, and this dynamics may manifest itself in change-point synchrocomplexes. The configuration of a change-point synchrocomplex (defined as the set of EEG channels containing the change-points which constitute a synchrocomplex) therefore can be an informative characteristic of the current brain state, and a set of such configurations found in a given EEG can represent the variety of brain states during the interval of the EEG recording. It is evident that such a set of configurations cannot be estimated only on the basis of the information provided by the pairwise analysis described above: the same mean values of the synchrony indices for the pairs of channels can be observed in the cases of absolutely different sets of the configurations of change-point synchrocomplexes. If the distances between neighbouring electrodes are roughly equal, the number of change-points participating in a synchrocomplex, which we call the order of synchrocomplex, provides an estimate of the total cortical area were the activity changes simultaneously.
An accurate definition of a change-point synchrocomplex must include a certain time threshold for change-points to be included, in a form of a short time window covering both the earliest and the latest change-points of the complex, or in a form of the maximal allowed time distance between successive change-points. In practice, a more crude way to define synchrocomplexes can be used for simplicity, especially if the probabilities of certain configurations should be computed: the analysed EEG recording is divided into fixed windows of equal lenght, and each window is characterized by the presence or absence of a change-point in each channel. The length of the "window" is chosen so that it covers no more than one change-point from each channel, and yet the cases when the border between two windows separate two apparently close (in time) change-points from different channels are not very frequent.
The probability of simultaneous appearing by chance of change-points in different channels falls very quickly with the increase of their number, so that, in practice, all the "higher order" synchrocomplexes can be considered as "real", showing some actual degree of connectiveness between EEG channels. For simplicity, we may consider only the number of synchrocomplexes of a given configuration as the main index, keeping in mind that the "low order" synchrocomplexes may result from casual coinciding of change-points and that they require special caution in the interpretation of results. The configurations of the synchrocomplexes can be ranged in accordance with their rate of occurence, and then only the most frequently found configurations are analysed.
The analysis of change-point synchrocomplexes can be illustrated more clearly if the spatial dimensionality of the EEG is reduced to one dimension by using the electrodes placed along one line. We applied the electrodes with equal distances between them as a "chain" from the forehead to the occipital (over the standard positions O2, P4, C4, F4, and between them). By this way we could not, of course, get the entire spatial picture of the cortical potentials, but, in return, gained in spatial resolution with relatively low number of electrodes (specifically, 16 electrodes).
Each of 10 artefact-free EEG recordings (1 minute, 7680 samples; eyes closed, resting) was subjected to the change-point detection and then was broken into 590 successive "windows" of 13 samples (about 100 ms) length. In 47.3% of "windows" no change-point was found in any of the channels. In 9.9% of "windows" a change-point was found only in one channel. The low number of such "singular" change-points can be attributed primarily to the dense positioning of the electrodes: the electric potential dynamics at close points on the scalp is very similar because of high functional connectivity of close cortical areas and because of volume conduction effects. The coincidence of change-points at different electrodes, if the spaces between them are not too large, is, however, very usual in more general situation (see, for example, Fig. 7.9).
The presence or absence of a change-point in a channel of a certain "window" can be denoted as 0 or 1, respectively. Then each window will be denoted by a combination of 16 binary digits, each digit corresponding to a channel. The combination describes the configuration of a synchrocomplex in this "window", if change-points are found in two or more channels simultaneously. A possible number of such combinations is rather high (216=65536) and it could not be convenient to analyse the rate of occurrence of each combination separately. In 10 minutes of our EEG recording (5900 "windows"), however, only 805 of the combinations were found, of which 788 described the configurations of synchrocomplexes, 16 -- "windows" with a change-point in only one channel and 1 -- no change-points in a "window" at all. The 788 configurations was still a large set, and it was important to see whether some of them were very rare and resulted from random coincidence of the change-points, or was unseparable from those "random" configurations.
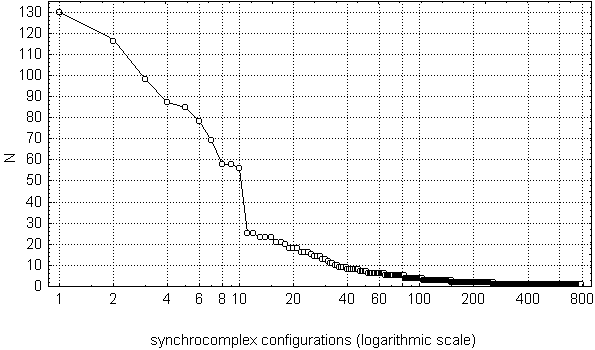
Fig. 7.13. Variation row of synchrocomplex configurations
Horizontal axis (logarithmic scale) represents synchrocomplex configurations
in occurrence rate decreasing order (each point on the axis corresponds to a
single configurations); vertical axes represents their occurrence rate (as
the total number of the synchrocomplexes with the given
configuration).
Subject EGL. 10 one-minute EEGs were recorded in eyes closing, resting condition.
Electrodes were placed with equal spaces between them along
anterior-posterior axis at right side of the head. Their
numbers started from the occipital. EEG was filtered with bandpass
7.5--12.5 Hz (alpha), the change-points detected in the basic diagnostic
sequence. Synchrocomplex configuration was defined by the
set of channels in which the coinciding change-points were found (for more
details see text).
As it can be seen from Fig. 7.13, most of the configurations were found very rarely: from 2 to 3 cases -- 152 configurations, 1 case -- 532 (!). Only 35 configurations were found more than 10 times (more rare than one per minute in average), but the total number of "windows" with these configurations was 1256 (21.3% of the total number of the "windows"), i.e., almost equal to the number of the other synchrocomplex configurations (1265, or 21.4%). Most prominent at the Fig. 7.13 are 10 most frequent configurations, each of which was found in no less than 56 "windows". Taken together they occupied about 1/7 of the total number of "windows", or 26.9% of those "windows" were at least one change-point was found.
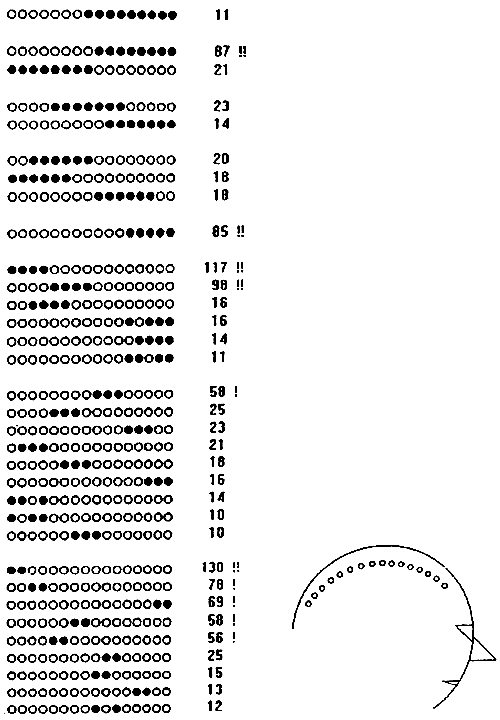
Fig. 7.14. Most frequent configurations of synchrocomplexes
Synchrocomplex configurations (change-points in alpha band power) found at least 10 times in 10 one-minute EEG recordings (same EEGs as in Fig. 7.13). Black circles: electrodes where change-points were detected. Electrode positions are shown in the right. N -- number of observations, "!" -- configurations with highest rate of change-point occurence.
Thus, a half of the synchrocomplexes found in these EEGs had one of 35 most frequent configurations, which is only 1/1872 of all possible configurations. These most frequent configurations are given in Fig. 7.14, and the top frequent 10 -- also in Fig. 7.15. Due to the linear scheme of electrode montage the linear notation used for synchrocomplex configurations in these tables give a picturial view of real spatial relations of the areas with synchronous changes of electric potential dynamics in alpha band on the head surface.
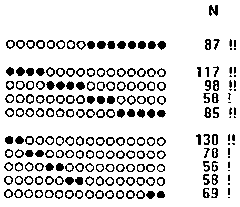
Fig. 7.15.
A remarkable characteristic of 30 of the 35 most frequent synchrocomplex configurations (given in Fig. 7.14) is their unbroken structure, i.e., all the channels with change-points in them were in succession. The five configurations with breaks were relatively infrequent (the most frequent of them, with change-points in channels 12,14,15 and 16, was found only 16 times during the 10 minutes of the EEG registration); in all these cases the break involved only one channel and the outer borders of these configurations exactly matched the outlines of the most frequently found (58 or more) synchrocomplexes. It is likely that the broken configurations were caused by artificial deviation from the frequent ones with a miss of a change-point. Thus, the areas under our electrode chain with simultaneously observed change-points in alpha band power typically were continuous (unbroken). This is an example of a fact which can be determined only with the multichannel, but not pairwise, analysis.
Another interesting spatial characteristic of these data was the higher rate of occurence of the higher order (i.e., more extended) synchrocomplexes in the anterior relative to posterior areas of the head. This phenomenon probably reflected the same intrinsic spatial features of the alpha activity as the higher level of change-point synchronization in anterior areas determined by the pairwise analysis.
The low percentage of the synchrocomplexes with broken configurations showed that the casual miss of a change-point only slightly affected the results. There were also another characteristic feature of the frequent configurations which could be observed only in the case of low influence of the missing change-points: the frequently found synchrocomplexes of the same or similar orders, as it can be seen in Table 7.2, did not overlapped. The method we used, though a very simplified, thus demonstrated a rather high ability to study the change-point synchrocomplexes.
We are now only in the beginning of the practical use of the EEG change-point synchrocomplex analysis, and it is too early to draw more certain conclusions on the basis of these data. The results described in this subsection should be considered not from the point of view of revealing certain physiological mechanisms but rather as an illustration of the potential of the multichannel approach to change-point synchronization analysis. In the next subsection we will discuss the general characteristics of the change-point syncronization methodology and give the outlines of some other ways of its developing.